第 六 章 鳥類與哺乳動物的起源
第 六 章
鳥類與哺乳動物的起源
根據進化論,生命的起源來自海洋,並且逐步進化成兩棲動物,然後遷移到陸地上的。這個學說認為,兩棲動物進化成爬行動物後,才形成了陸地動物。在生理學與解剖學中,如此的進化是不可能的。例如,在水中孵化的兩棲動物的卵,逐步進化成在乾燥環境下孵化的爬行動物的卵,其沒有可能性的證據大量存在。當我們注意地質記錄時就可以發現,這樣的進化是不存在的:爬行動物與無脊椎動物之間毫無聯繫,也不是以同一“祖先”出現在地球上的。在脊椎動物古生物學方面,就連權威的進化論者、古生物學家羅伯特•L•卡羅爾也不得不接受“早先的爬行動物與兩棲動物有著極大的區別,並且至今仍未發現它們的‘祖先’。”44
然而,進化論者註定沒有希望的童話故事,依然沒有結束。他們還面臨著讓陸生動物“飛翔”的難題呢!進化論者相信,鳥類必須是以一定的形式進化而成的,因此,他們認定鳥類是從爬行動物“改造”而來的。但是,陸地動物與鳥類有著完全不同的身體結構,這種逐漸進化而來的解釋也是不合理的。首先,鳥的顯著特性——翅膀,僅僅這一點就使進化論者面臨很大的困境。土耳其進化論者安金•柯儒(Engin Korur)認為,翅膀沒有進化的可能性:“眼睛和翅膀的共同特性在於,只有在它們完全發育了的時候,才能起到有價值的作用。換句話說,殘缺發育的眼睛是看不見的,單翅鳥的翅膀是飛不起來的。至於這些器官是怎樣形成的,仍然是需要說明的自然秘密之一。”45
正如我們所見,結構完美的翅膀怎樣形成的這一問題,用連續的偶然突變而形成的解釋,是完全回答不了的;根本沒有辦法解釋,爬行動物的前臂怎麽能在基因不破壞(突變)的情況下,形成了完美的雙翼。而且,進化成鳥類的陸地動物,僅僅擁有翅膀是不夠的;陸生動物缺乏鳥類用於飛翔的其他結構性機制。例如,鳥類的骨頭比陸生動物的輕得多;還有,它們的肺功能是非常不同的;它們有不同的肌肉和骨骼系統,以及非常專業化的心臟循環系統。所有這些機制必須同時並完整的存在,它們不會逐漸通過“積累”來形成。這就斷定陸生動物向飛行動物的進化,是一個荒謬的理論。這還帶來了另一個問題:即使我們認為這種不科學的假想是真的,那麽,為什麽還是不能發現一些“半翼”或“單翼”的飛行動物的生物化石,來支持他們的假想呢?從解剖學來看,鳥類和爬行動物有很大的區別。
鳥類特殊的肺
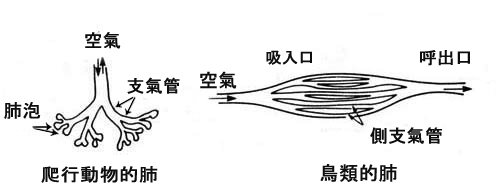
鳥類的肺系統和爬行動物的肺系統完全相反。陸生動物都是用同一進氣管道來呼吸的,而鳥類從肺的前面吸氣、從肺的後面呼氣。這種系統是為鳥類在高空飛行時,對氧氣的需要增多而特別“設計”的;這種設計依靠陸生動物的進化是不可能形成的。因為,這兩種肺系統的過渡形式,不可能具備呼吸功能。
另一所謂的過渡形式:始祖鳥
面對為什麽找不到“單翼”、“半翼”的動物化石的問題,進化論者們提出了一種特別的動物,他們仍然聲稱那是少數過渡形式中最有名的、叫做“始祖鳥”的鳥類化石。進化論者認為始祖鳥是現代鳥的祖先,它們生活在大約1.5億年以前。其理論認為,在一些名為Velociraptor或者Dromeosaur的小體形恐龍中,有一部分通過進化獲得了翅膀,然後開始飛行。始祖鳥被認為是從恐龍祖先中分離出來,並且開始飛行的第一個生物。進化論者很快把這個故事廣泛地宣傳起來。但是,從對始祖鳥化石的最新研究來看,證明這種動物絕對不是一個過渡形式,而是一種與現代鳥類,有些不同特性的鳥類罷了。至於始祖鳥是不能完全飛行的“半鳥”的觀點,直到最近,進化論者仍不斷提到它,而且認為這個生物的胸腔內缺乏胸骨,這便是它不能持續飛行的最重要的證據。(胸骨是飛行時用來固定胸腔肌肉的一塊骨頭。現今,在會飛行和不會飛行的鳥類中,甚至與鳥類完全不同的哺乳動物蝙蝠,都擁有這樣的胸骨。)
可是,在1992年發現的第七塊始祖鳥的化石,在進化論者中間引起了巨大的震動。因為,在這塊新發現的始祖鳥的化石中,進化論者長時期認為並不存在的胸骨,實際上卻存在著。
《自然》雜誌對這塊化石的描述如下:
“最近發現的第七塊始祖鳥化石,顯示了長期以來所懷疑的、然而從未得到證明的一塊矩形胸骨的存在。這證明它具有強壯的飛行肌肉。”46
這個發現使始祖鳥屬於不能完全飛行的“半鳥”的說法無效了。
另一方面,始祖鳥作為真正意義上的鳥的最重要的證據之一,就是鳥類的羽毛結構;始祖鳥與現代鳥毫無區別的非對稱的羽毛結構,表明了始祖鳥能完美地飛行。正如著名的古生物學家卡爾•O•鄧巴所說的:“就因為它的羽毛,從中可見始祖鳥顯然屬於真正的鳥類。”47
始祖鳥的羽毛還揭示了另一個事實,那就是該鳥溫血性的新陳代謝。眾所周知,爬蟲動物和恐龍是隨環境溫度而影響其體溫的冷血動物,而不是獨立調節它們體溫的。而鳥類羽毛的非常重要的功能之一,就是保持體溫。始祖鳥有羽毛的事實顯示,它是需要保持體溫的真正的溫血動物;這與恐龍形成了對比。
進化論者的假想:始祖鳥的牙齒和爪子
進化論者把始祖鳥當作一個過渡形式的兩個要點是:它的牙齒和爪子。是的,始祖鳥的嘴裏有牙齒,翅膀上有爪子。但是,這些特性並不意味著它與爬行動物之間,有任何一種聯繫。而且,當今存在的兩種鳥類:焦鵑(Touraco)和麝雉(Hoatzin)都有可以抓住樹枝的兩個爪子;它們是完全沒有爬行動物特性的鳥。就憑始祖鳥的翅膀上有爪子,而認定它是過渡形式的主張是無效的;同樣,憑藉始祖鳥嘴裏的牙齒,而認定它是過渡形式也是無效的。進化論者利用這些牙齒,證明始祖鳥具有爬行動物的特性,是有意識的欺騙。況且,牙齒並不是爬行動物的一種典型特性。今天,一些爬行動物有牙齒,另一些則沒有牙齒。更重要的一點是,始祖鳥不是有牙齒的唯一鳥類。我們可以確信,有牙齒的鳥類今天已不存在,但從化石記錄看,可以發現與始祖鳥相同及以後的時期內,甚至與我們更近的時期裏,有過這樣一種特殊的鳥種,人們將之分類為“齒鳥”。
最重要的一點是,始祖鳥和其他齒鳥的牙齒結構,完全不同于被說成是它們祖先的恐龍的牙齒。根據著名鳥類學家馬丁(Martin)、施締沃特(Steward)和威茨同(Whetstone)的觀測,始祖鳥和其他齒鳥的牙齒上端是平整的,而且有寬大的牙根。然而,進化論者當成這些鳥的祖先的獸腳亞目食肉恐龍(theropod)的牙齒,卻像鋸一樣的突出,而且牙根狹窄。48
研究人員也比較了始祖鳥和當成它們祖先的恐龍的腕骨,結果並沒有發現它們之間有任何相似之處。49
約翰•奧斯特羅姆,這位聲稱始祖鳥從恐龍逐步進化而來的權威,根據解剖學家塔爾斯坦諾(Tarsitano)、赫克特(Hecht)、A•D•沃克(A.D.Walker)的研究,指出那些存在於這種動物與恐龍之間的“相象”,實際上是錯誤的解釋。50
所有這些結論表明,始祖鳥不是過渡形式,而是只能分類為“齒鳥”的一種鳥類。
始祖鳥與其他古老鳥類的化石
當進化論者花費數十年時間,根據始祖鳥與其他古老鳥類的化石,指出那是始祖鳥從其他古鳥進化而來的最大證據時,而近期發現的化石卻從其他方面使這個假想變得無效。
鳥毛的設計
就鳥的骨骼結構而言,鳥類的肺系統和常溫下的新陳代謝,都不同於爬行動物。鳥類與爬行動物之間另一巨大的差別,就是鳥類具有完美形式的羽毛結構,爬行動物的身體覆蓋著鱗片,而鳥的身體上長滿了羽毛。
進化論者考慮爬行動物是鳥的祖先的同時,又聲稱鳥類的羽毛是從爬行動物的鱗片上進化而來的。但是,鱗片和羽毛之間並沒有任何相象之處。
美國康涅狄格大學的神經生物學、生理學教授,A•H•布拉士,儘管他是個進化論者,但仍接受這個現實:"就發育而言,每一羽毛的基因結構和組織,外貌形成與軟組織都是不相同的。"1 而且,布拉士教授檢查鳥毛的蛋白質結構後爭議道,那是"脊椎動物所獨有的"特徵。2
沒有化石證據表明,鳥毛是從爬行動物的鱗片進化而來的。相反,如布拉士教授所說的:"在地質記錄裏,羽毛是突然出現的——作為區別鳥類特徵的'不可否認的獨特'出現的。"3 而且,在爬行類動物中,並沒有發覺它們的表皮結構給鳥類提供羽毛的根源。4 1996年,古生物學家製造了一個"所謂有羽毛的恐龍的化石在中國被發現"的故事,並稱之為"中國始祖鳥"。1997年,人們發現,這些化石事實上與鳥的羽毛結構毫無相似之處。5
另一方面,當我們仔細檢查鳥的羽毛時,發現了用進化論無法解釋的複雜結構。著名鳥類學家艾倫•費都克斯雅(Feduccia)說明:"羽毛具有空氣動力學的功能。它們重量極輕,有足夠的緩慢提速的飛行能力,並且非常容易地恢復到原有的位置。"他在錯誤的進化論學說面前承認:"我無法理解為適應飛翔而設計的這個器官,怎樣會在當初由另一個功能轉變而來。"6
羽毛的設計也使查理斯•達爾文不得不考慮它們。而且,孔雀羽毛奇妙的美學形態已使他的理論"得病了"(他自己的話)。1860年4月3日,他給朋友阿薩•格瑞的信中寫道:"我記得很清楚,在考慮眼睛的理論時,我渾身發冷,但我已經熬過了這個階段……"。他繼續說:"……現在,一些不重要的生物結構性方面的細節,使我經常感到不舒服。孔雀尾巴上的那些羽毛,我每每注視它們時,使我得病了!"7
這就是進化論無法解釋的奇妙設計——鳥的羽毛。
1,A.H.布拉士著"羽毛的起源",《進化生物學》雜誌1996第9卷,第132頁。
2,同上,第131頁。
3,同上,第133頁。
4,同上,第131頁。
5,"採集有羽毛的恐龍",《科學》雜誌第278卷1229頁,1997年11月14日。
6,道格拉斯•帕默爾《學會飛翔》("評艾倫•費都克斯雅《鳥的起源與進化》",耶魯大學出版社,1996年)《新科學家》雜誌第153卷第44頁,1997年3月1日。
7,諾曼•麥克佩斯《呼喚理性:重審達爾文》的"開場白",第101頁。波士頓,1971年。

"孔子鳥"與始祖鳥同齡。

仔細察看鳥的羽毛,
就會發現那是數千根細毛彼此相連的。
這種獨特的設計基於空氣動力學原理。
1995年,中國脊椎古生物學研究所的兩名古生物學家侯聯海(Lianhai Hou)、周中和(Zhonghe Zhou)發現了一塊新的鳥化石,他們把它命名為“孔子鳥”。這只鳥幾乎與始祖鳥同齡(約1.4億歲左右),但它的嘴裏根本沒有牙齒。還有,它有著與今天的鳥兒一樣的嘴巴和羽毛,有著與現代鳥一樣的骨骼,翅膀上還有與始祖鳥一樣的爪子。被稱為“尾綜骨”的特殊結構,支撐這種鳥的尾毛。一句話,這種與始祖鳥同齡的鳥(被認為是所有鳥類最古老的祖先,並稱之為“半爬行動物”),與現代的鳥很相似。這個事實使進化論關於始祖鳥,是所有鳥類祖先的說法無效了。51

1996年11月,在中國發掘出另一塊化石,這引起了更大的混亂。侯(Hou)、馬丁和艾倫•費都加(Feduccia)在《科學》雜誌上宣佈,發現了一個命名為“遼寧鳥”、有著1.3億年歲的“老傢伙”。 遼寧鳥有一塊可以支撐飛行肌肉的胸骨——與現代鳥一樣。這只鳥在其他方面與現代鳥也沒有區別。唯一不同的是嘴裏的牙齒。這表明這種有牙齒的鳥,根本沒有像進化論者所聲稱的那樣,有一個原始結構。52就像艾倫•費都加在《發現》(Discover)雜誌裏解釋的那樣:“這些(遼寧)鳥來自何處?其化石說明它並非來自恐龍。”53
另一個使進化論者關於始祖鳥的主張崩潰的化石是Eoalulavis。Eoalulavis據說比始祖鳥年輕3千萬歲,約有1,2億年,其翅膀結構與當今的一些飛鳥相似,飛起來緩慢。這證明,在1.2億年前,它們和現代鳥在很多方面沒有什麽區別,同樣在空中飛行。54
這些事實再次證明,始祖鳥和與其相似的別的古老鳥類,決不是什麽過渡形式。化石沒有顯示不同的鳥類會相互進化。相反,地質記錄證明,今天的鳥類和諸如始祖鳥的原始鳥類,實際上是同時共存的。但是,其中的一些鳥類,像始祖鳥和孔子鳥都已經滅絕了,僅有一些先存的種類能夠延續至今。
總之,始祖鳥的某些特別的特徵,無法說明它是一種過渡形式的鳥!
事實上,哈佛大學的兩個有名的進化論家、古生物學家斯帝芬•簡•古爾德和奈爾斯•埃爾德雷奇也接受:始祖鳥是形體上具有各種不同特徵的“鑲嵌式的”生命體,但它永遠不能被看作是一個過渡形式。55
想像之鳥——與恐龍的聯繫
進化論者試圖斷定,始祖鳥是一種過渡形式的鳥,進而宣稱鳥是從恐龍進化來的。但是,世界上最著名的鳥類學家之一、北卡羅來納大學的艾倫•費都加,儘管他自己是個進化論者,可反對鳥與恐龍有親緣關係的理論。費都加就此指出:
“我研究鳥的頭顱已經25年了,然而,我沒有看出它們之間有任何相象之處,我只是看不見它們……。在我看來,獸腳亞目食肉恐龍是鳥的起源的理論,這將是二十世紀古生物學中最大的尷尬。”56
拉裏•馬丁,堪薩斯大學的古代鳥類專家,也反對鳥類與恐龍具有相同血統的理論。在討論這一相互矛盾的進化論時,馬丁說:
“告訴你們實情吧,如果我必須支援具有恐龍特徵的鳥類起源於恐龍的話,每次我都陷入尷尬中,並且不得不站起來為之辯解。”57
總之,只依賴於始祖鳥而豎立的“鳥的進化”的假想,僅僅是懷有偏見者和進化論者想像力的產物。
哺乳動物的起源
正如我們前面所陳述的那樣,進化論試圖把一些想像中的海洋生物轉變成爬行動物,把鳥類也說成由爬行動物進化而來。按照進化論相同的假想,認為爬行動物不僅是鳥,而且還是哺乳動物的祖先。但是,爬行動物之間存在大的結構性差異;它們的身上有鱗片、屬於冷血動物,並且通過產卵繁殖後代;而哺乳動物的身上有皮毛、屬溫血動物、通過直接生育活生生的後代來繁衍的。
蒼蠅的起源是什麽?
圖片下方文字] 進化論假想的一個例子:恐龍捕捉蒼蠅時突然展翅起飛的。.

進化論者聲稱鳥類由恐龍進化而來的同時,又這樣支持他們的斷言:一些恐龍撲打著前腿去捕捉蒼蠅,如圖所示的“展翅飛翔”。
這種毫無科學基礎與十足的想像,還存在簡單的邏輯上的矛盾,那就是進化論者解釋蒼蠅起源的例子,即蒼蠅是已經擁有完美飛行能力的物種。
人類的眼睛不能在一秒鐘開合10次,而一隻蒼蠅能在一秒鐘平均拍打其翅膀達500次。而且,蒼蠅同時移動它的雙翅;翅膀在振動過程中最輕微的不協調,將使蒼蠅失去平衡。但是,這樣的事從未發生。進化論者首先應該解釋,蒼蠅如何獲得飛行能力。甚至家蠅完美的造化形式,使進化論的假想無效了。
英國生物學家羅賓•伍頓在一篇題為“蒼蠅翅膀的機械設計”的文章裏寫道:“我們更好地理解昆蟲翅膀的作用時,更為精細和美麗的設計就顯露了。結構的設計在傳統上盡可能少地使其變形;機械設計也是便於用預設的方式移動其元件的。”
昆蟲的翅膀把上面提到的構造合為一體:使用具有廣角彈性的組成部分的雙翼,這些高雅的組合允許適當的變形,以便適應外界阻力,並最大可能地利用空氣。尚無任何技術上與蒼蠅的翅膀相似的設計。1
另一方面,也沒有哪怕一塊能證明蒼蠅想像中進化而來的化石。法國動物學家格拉塞(Grassé)因之說道:“關於昆蟲的起源,我們仍然處在黑暗之中。”2
2,格來塞:《有生物的進化》第30頁,紐約學術出版社,1977年。

進化論者認為,所有的哺乳動物來自同一個祖先,但是象熊、鯨、鼠及蝙蝠等動物之間卻有著很大的區別,它們都有各自不同的特殊結構。例如,蝙蝠具有在黑夜裏辯明方向的、非常敏感的聲波系統;連現代技術都無法設計出如此複雜的系統。進化論者卻聲稱這是偶然形成的;這是不可能的事。事實上,地質記錄顯示,現代蝙蝠的完美結構突然產生,並沒有任何進化的跡象。

5000萬年的蝙蝠化石與現代的蝙蝠毫無兩樣。(《科學》雜誌154卷)
爬行動物和哺乳動物結構性的差異,在於它們的顎結構上。哺乳動物的下顎只由一塊顎骨組成,牙齒就成長在這塊骨頭上;在爬行動物中,其下顎的兩邊各有三塊小骨。另一個基本的差別是,所有哺乳動物的中耳內有三塊骨(砧骨、錘骨和鐙骨)。在所有的爬行動物中,它們的中耳內只有一塊骨。進化論者聲稱,爬行動物的顎和耳,逐步進化成了哺乳動物的顎和耳。然而,對這種變化怎樣發生的問題,依然無法給予答復。單骨的耳朵怎麽逐步進化到三骨的耳朵?在此期間聽力怎樣起作用?這依然是他們永遠不能回答的難題。這並不奇怪,因為連接爬行動物和哺乳動物之間過渡形式的一塊化石都沒有發現過。因此,進化論古生物學家羅傑•盧因被迫說道:“第一個哺乳動物的過渡仍然是個謎,它大概經過一個或至多兩個世系才會發生。”58
在20世紀,進化論權威、新達爾文學說的創始人之一喬治•蓋洛德•辛普森,針對進化論者困惑的事實做了如下評論:
“在地球上生命歷史的長河中,最讓人迷惑的事情,來自中生代的變化,即爬行動物時代向哺乳動物時代的突然轉變。這好象原本由爬行動物唱主角的舞臺突然謝幕了,尤其是數量龐大、種類繁多的恐龍,立即站起來再次搭起了場地,只不過演員變成了哺乳動物——恐龍根本沒有出現的新角色;別的爬行動物被推到一邊,所有主角由早先‘演過戲’的不同種類的哺乳動物出演了。一個新的時代開始了。”59
這個出現的哺乳動物,好象在過去的時代裏從未出現過似的。此外,當哺乳動物突然出現時,他們彼此互不相同。像蝙蝠、馬、老鼠和鯨魚等全都是哺乳動物,並且在相同的地質時期出現。不論使用多麽奇異的想像力,在它們之間建立進化關係也是不可能的。
進化論者、動物學家埃裏•克朗伯特在《進化》雜誌裏發表的一篇文章這樣寫道:
“為了在哺乳動物的族類裏建立進化的親緣關係(解剖學的鏈結),而尋找資訊的人們將會感到失望。”60
這一切證明,所有的生物都是突然出現並且完全形成的,其間沒有任何進化的過程。這是它們被創造的具體證據。但進化論者們卻努力解釋說,生物物種作為進化的跡象,以特別的順序產生的。然而,生命體出現的順序是“創造的順序”,因為根本談不上進化的過程。化石記錄顯示:通過完美無缺的創造,地球上先是海洋、然後是充滿生命體的陸地,直至最後創造了人類。與“南方巧猿”的故事恰恰相反——他們憑藉強大的媒體渲染,強迫人們接受人類是從猿變來的。而人類在地球上也是突然出現、完整形成的。
馬的進化的神話

直到最近,按照一個虛構的順序推測,把馬的進化作為進化論學說主要的化石證據。但是,今天很多進化論者自己坦白地承認,馬的進化的假想已經破產。1980年,在芝加哥的菲爾德自然博物館召開了為期四天的專題座談會,有150位進化論者參加了這次會議。會議就沒有化石記錄、也沒有逐漸進化記載的假想作了討論——對馬的進化的假想。進化論者博伊斯•仁斯柏格(Rensberger)在會上說:“馬的進化的例子很流行:是由差不多5000萬年以前就有的四趾狐狸型生物,按照漸進順序,進化成今天比狐狸大得多的一趾蹄的馬;這早已是眾所周知的錯誤。與所謂逐漸變化相反,每個過渡形式的化石的出現,是截然不同、始終不變的,隨後絕種以及過渡的形式都是未知的。1
仁斯柏格發表針對馬的進化假想的誠實觀點的同時,事實上也提出了該學說在地質記錄中所面臨的最大困境“過渡形式”。著名古生物學家、英國自然史博物館主任科林•派特森,在博物館的底層仍然給公眾看的關於“馬的進化”展覽會上說:“在究竟何謂那段生命(馬的進化史——譯注)的歷史方面,有太多的故事,其中一些比其他更具想像力。最著名的例子,就是仍在樓下展覽的、或許在50年以前就已經準備好的關於馬進化的展品。馬的進化已經作為真理不斷寫入教科書中。現在,我認為這種做法是不愉快的,尤其是提出這些故事的人們,他們自己也意識到這些材料中有投機的成分。”2 那麽,“馬的進化”的假想的基礎是什麽?進化論者用豐富的想像力,通過對生活在印度、北美洲、南美洲及歐洲不同時期不同生物化石有序地排列,闡釋了這一騙人的假想圖表;由不同研究人員提供的、馬的進化的圖表達20多張。對這些彼此不同的譜系圖,進化論者之間並沒有達成共識。其中唯一的共同點是,5500萬年前生活在始新世時期的、類似於狗的一種被稱為“始祖馬”的動物,是馬的最初祖先。但是,這種虛構的從始祖馬到馬的進化聯繫,是完全矛盾的。這種馬與至今生活在非洲、甚至與馬沒有一點聯繫與相似之處的“蹄兔”是同類動物。3
有關馬的進化的毫無根據的斷言,從不斷發現的地質記錄中破滅了;始祖馬與現代馬種(西方馬種和內華達州馬種)的化石顯示,現代馬和他們所稱的馬的祖先,是在同一時期生存的物種。這清楚地說明馬的進化在任何時候都不存在。4
進化論者、作家戈登•R•泰勒,在《大進化秘密》一書中,對達爾文缺乏根據的觀點解釋道:“也許,達爾文學說最大的弱點,是古生物學家沒能發現令人信服、演示生物體主要進化性變化的發育與順序的證據……。馬是經常用來解決這一問題的唯一例子。但是,實際上始祖馬與馬類的聯繫是不可思義的;他們聲稱馬的體型連續增加著,可一些變化顯示,它們卻比始祖馬變小了,而不是變大了。把不同來源的標本放到一起,就構成看似具有說服力的進化順序,但實際上並不存在按時、按序排列的證據。5
這些雄辯的事實證明,達爾文學說作為牢固的證據,來描繪馬進化的圖表,不過是荒謬和難以置信的故事而已。
2,科林•派特森,“哈珀”,1984年2月,第60頁
3,法蘭西斯•赫廷,《長頸鹿的脖子:達爾文誤入歧途》,紐約Tick nor and Fields,1982年,第30-31頁
4,法蘭西斯•赫廷,《長頸鹿的脖子:達爾文誤入歧途》,第30-31頁
5,戈登•拉特雷•泰勒,《大進化秘密》,倫敦,天體書第230頁,1984年]
笔记
44 Robert L. Carroll, Vertebrate Paleontology and Evolution, New York: W. H. Freeman and Co., 1988, p. 198. ![]()
45 Engin Korur, "Gözlerin ve Kanatlar›n S›rr›"(The Mystery of the Eyes and the Wings), Bilim ve Teknik, No. 203, October 1984, p. 25. ![]()
46 Nature, Vol 382, August, 1, 1996, p. 401. ![]()
47 Carl O. Dunbar, Historical Geology, New York: John Wiley and Sons, 1961, p. 310. ![]()
48 L. D. Martin, J. D. Stewart, K. N. Whetstone, The Auk, Vol 98, 1980, p. 86. ![]()
49 Ibid, p. 86; L. D. Martin "Origins of Higher Groups of Tetrapods", Ithaca, New York: Comstock Publising Association, 1991, pp.485, 540. ![]()
50 S. Tarsitano, M. K. Hecht, Zoological Journal of the Linnaean Society, Vol 69, 1985, p. 178; A. D. Walker, Geological Magazine, Vol 177,1980, p. 595. ![]()
51 Pat Shipman, "Birds do it... Did Dinosaurs?", New Scientist, February 1, 1997, p. 31. ![]()
52 "Old Bird", Discover, March 21, 1997. ![]()
53 Ibid. ![]()
54 Pat Shipman, "Birds Do It... Did Dinosaurs?", p. 28. ![]()
55 S. J. Gould & N. Eldredge, Paleobiology, Vol 3,1977, p. 147. ![]()
56 Pat Shipman, "Birds Do It... Did Dinosaurs?", p. 28. ![]()
57 Ibid. ![]()
58 Roger Lewin, "Bones of Mammals, Ancestors Fleshed Out", Science, vol 212, June 26, 1981, p. 1492. ![]()
59 George Gaylord Simpson, Life Before Man, New York: Time-Life Books, 1972, p. 42. ![]()
60 R. Eric Lombard, "Review of Evolutionary Principles of the Mammalian Middle Ear, Gerald Fleischer", Evolution, Vol 33, December 1979, p. 1230. ![]()